Почему модель хищник жертва является не точной. Равновесие хищник—жертва
Часто представители одного вида (популяции) питаются представителями другого вида.
Модель Лотки – Вольтерры – модель взаимного существования двух популяций типа «хищник – жертва».
Впервые модель «хищник – жертва» была получена А. Лоткой в 1925 году, который использовал ее для описания динамики взаимодействующих биологических популяций. В 1926 году независимо от Лотки аналогичные (к тому же более сложные) модели были разработаны итальянским математиком В. Вольтерра, глубокие исследования которого в области экологических проблем заложили фундамент математической теории биологических сообществ или т. н. математической экологии.
В математической форме предложенная система уравнений имеет вид:
где x – количество жертв, y – количество хищников, t – время, α, β, γ, δ – коэффициенты, которые отражают взаимодействия между популяциями.
Постановка задачи
Рассмотрим закрытое пространство, в котором существуют две популяции – травоядные («жертвы») и хищники. Считается, что животных не ввозят и не вывозят и что еды для травоядных животных достаточно. Тогда уравнение изменения числа жертв (только жертв) примет вид:
где $α$ – коэффициент рождаемости жертв,
$x$– размер популяции жертв,
$\frac{dx}{dt}$ – скорость прироста популяции жертв.
Когда хищники не охотятся, они могут вымирать, значит, уравнение для количества хищников (только хищников) примет вид:
Где $γ$ – коэффициент убыли хищников,
$y$ – размер популяции хищников,
$\frac{dy}{dt}$ – скорость прироста популяции хищников.
При встрече хищников и жертв (частота встреч прямо пропорциональна произведению) хищники уничтожают жертв с коэффициентом, сытые хищники могут воспроизводить потомство с коэффициентом. Таким образом, система уравнений модели примет вид:
Решение задачи
Построим математическую модель совместного существования двух биологических популяций типа «хищник – жертва».
Пусть две биологические популяции совместно обитают в изолированной среде. Среда является стационарной и обеспечивает в неограниченном количестве всем необходимым для жизни один из видов – жертвы. Другой вид – хищник – также обитает в стационарных условиях, но питается только жертвами. В роли хищников могут выступать коты, волки, щуки, лисы, а в роли жертв – куры, зайцы, караси, мыши соответственно.
Для определенности рассмотрим в роли хищников – котов, а в роли жертв – кур.
Итак, куры и коты живут в некотором изолированном пространстве – хозяйственном дворе. Среда предоставляет курам питание в неограниченном количестве, а коты питаются только курами. Обозначим через
$х$ – количество кур,
$у$ – количество котов.
Со временем количество кур и котов меняется, но будем считать $х$ и $у$ непрерывными функциями от времени t. Назовем пару чисел $х, у)$ состоянием модели.
Найдем каким образом изменяется состояние модели $(х, у).$
Рассмотрим $\frac{dx}{dt}$ – скорость изменения количества кур.
Если котов нет, то количество кур возрастает и тем быстрее, чем больше кур. Будем считать зависимость линейной:
$\frac{dx}{dt} a_1 x$,
$a_1$ – коэффициент, который зависит только от условий жизни кур, их естественной смертности и рождаемости.
$\frac{dy}{dt}$ – скорость изменения количества котов (если нет кур), зависит от количества котов y.
Если кур нет, то количество котов уменьшается (у них нет пищи) и они вымирают. Будем считать зависимость линейной:
$\frac{dy}{dt} - a_2 y$.
В экосистеме скорость изменения количества каждого вида также будем считать пропорциональным его количеству, но только с коэффициентом, зависящим от количества особей другого вида. Так, для кур этот коэффициент уменьшается с увеличением количества котов, а для котов возрастает с увеличением количества кур. Будем считать зависимость также линейной. Тогда получим систему дифференциальных уравнений:
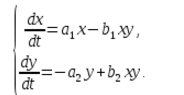
Данная система уравнений называется моделью Вольтерра-Лотки.
a1, a2, b1, b2 – числовые коэффициенты, которые называют параметрами модели.
Как видно, характер изменения состояния модели (x, y) определяется значениями параметров. Изменяя данные параметры и решая систему уравнений модели, можно исследовать закономерности изменения состояния экологической системы.
С помощью программы MATLAB система уравнений Лотки-Вольтерра решается следующим образом:
На рис. 1 представлено решение системы. В зависимости от начальных условий решения разные, чему отвечают разные цвета траекторий.
На рис. 2 представлены те же решения, но с учетом оси времени t (т.е. наблюдается зависимость от времени).
Взаимодействие особей в системе «хищник - жертва»
Студента 5 курса 51 А группы
отделения Биоэкологии
Назарова А. А.
Научный руководитель:
Подшивалов А. А.
Оренбург 2011
ВВЕДЕНИЕ
ВВЕДЕНИЕ
В своих ежедневных рассуждениях и наблюдениях мы, сами того не зная, а часто даже не осознавая, руководствуемся законами и идеями, открытыми много десятилетий назад. Рассматривая проблему хищник – жертва, мы догадываемся, что опосредованно жертва тоже влияет на хищника. Чем бы обедал лев, если бы не было антилоп; что бы делали управленцы, если бы не было рабочих; как развивать бизнес, если у покупателей нет средств…
Система «хищник-жертва» - сложная экосистема, для которой реализованы долговременные отношения между видами хищника и жертвы, типичный пример коэволюции. Отношения между хищниками и их жертвами развиваются циклически, являясь иллюстрацией нейтрального равновесия.
Изучение данной формы межвидовых взаимоотношений, помимо получения интересных научных результатов, позволяет решать многие практические задачи:
оптимизация биотехнических мероприятий как по отношению к видам-жертвам, так и по отношению к хищникам;
улучшение качества территориальной охраны;
регуляция прессинга охоты в охотхозяйствах и т. д.
Выше сказанное определяет актуальность выбранной темы.
Целью курсовой работы является изучение взаимодействия особей в системе «хищник - жертва». Для достижения цели поставлены следующие задачи:
хищничество и его роль в формировании трофических взамоотношений;
основные модели взаимоотношения «хищник - жертва»;
влияние общественного образа жизни в стабильности системы «хищник - жертва»;
лабораторное моделирование системы «хищник - жертва».
Совершенно очевидно влияние хищников на численность жертв и наоборот, однако определить механизм и сущность этого взаимодействия достаточно сложно. Эти вопросы я намерен раскрыть в курсовой работе.
#�������################################################"#5#@#?#8#;#0###��####################+###########��\############### ###############��#���############# Глава 4ГЛАВА 4. ЛАБОРАТОРНОЕ МОДЕЛИРОВАНИЕ СИСТЕМЫ «ХИЩНИК - ЖЕРТВА»
Ученые университета Дьюка в сотрудничестве с коллегами из Стэндфордского университета, Медицинского института Говарда Хьюза и Калифорнийского технологического института, работающие под руководством доктора Линчона Ю (Lingchong You), разработали живую систему из генетически модифицированных бактерий, которая позволит более детально изучить взаимодействия хищника и жертвы на популяционном уровне.
Новая экспериментальная модель является примером искусственной экосистемы, для создания которой исследователи программируют бактерии на выполнение новых функций. Такие перепрограммированные бактерии могут найти широкое применение в медицине, очистке окружающей среды и создании биокомпьютеров. В рамках данной работы ученые переписали «программное обеспечение» кишечной палочки (Escherichia coli) таким образом, что две разных бактериальных популяции сформировали в лабораторных условиях типичную систему взаимодействий хищник-жертва, особенностью которой являлось то, что бактерии не пожирали друг друга, а управляли численностью популяции-оппонента посредством изменения частоты «самоубийств».
Направление исследований, известное как синтетическая биология, возникло примерно в 2000 году, и в основе большинства созданных с тех пор систем лежит перепрограммирование одной бактерии. Разработанная авторами модель уникальна тем, что он состоит из двух живущих в одной экосистеме бактериальных популяций, выживание которых зависит друг от друга.
Ключевым моментом успешного функционирования такой системы является способность двух популяций взаимодействовать между собой. Авторы создали два штамма бактерий – «хищников» и «травоядных», в зависимости от ситуации высвобождающими в общую экосистему токсичные либо защитные соединения.
Принцип действия системы основан на поддержании соотношения количества хищников и жертв в регулируемой среде. Изменения количества клеток одной из популяций активируют перепрограммированные гены, что запускает синтез определенных химических соединений.
Так, малое количество жертв в среде вызывает активацию гена самоуничтожения в клетках хищника и их гибель. Однако, по мере увеличения численности жертв, высвобождаемое ими в среду соединение достигает критической концентрации и активирует ген хищника, обеспечивающий синтез «антидота» к суицидальному гену. Это ведет к росту популяции хищников, что, в свою очередь, приводит к накоплению в среде синтезируемого хищниками соединения, толкающего жертв на самоубийство.
С помощью флуоресцентной микроскопии ученые документировали взаимодействия между хищниками и жертвами.
Клетки-хищники, окрашенные в зеленый цвет, вызывают самоубийство клеток-жертв, окрашенных красным. Удлинение и разрыв клетки-жертвы свидетельствует о ее гибели.
Эта система не является точным отображением взаимодействий хищник-жертва в природе, т.к. бактерии-хищники не питаются бактериями-жертвами и обе популяции конкурируют за одни и те же пищевые ресурсы. Однако авторы считают, что разработанная ими система является полезным инструментом для биологических исследований.
Новая система демонстрирует четкую взаимосвязь между генетикой и динамикой популяций, что в будущем поможет при изучении влияния молекулярных взаимодействий на популяционные изменения, являющиеся центральной темой экологии. Система предоставляет практически неограниченные возможности изменения переменных для детального изучения взаимодействий между окружающей средой, регуляцией генов и популяционной динамикой.
Таким образом, с помощью управления генетического аппарата бактерии позволяет имитировать процессы развития и взаимодействия более сложных организмов.
ГЛАВА 3
ГЛАВА 3. ВЛИЯНИЕ ОБЩЕСТВЕННОГО ОБРАЗА ЖИЗНИ В СТАБИЛЬНОСТИ СИСТЕМЫ «ХИЩНИК - ЖЕРТВА»
Экологи из США и Канады показали, что групповой образ жизни хищников и их жертв радикально меняет поведение системы «хищник–жертва» и придает ей повышенную устойчивость. В основе данного эффекта, подтвержденного наблюдениями за динамикой численности львов и антилоп гну в парке Серенгети, лежит то простейшее обстоятельство, что при групповом образе жизни снижается частота случайных встреч хищников с потенциальными жертвами.
Экологи разработали целый ряд математических моделей, описывающих поведение системы «хищник–жертва». Эти модели, в частности, хорошо объясняют наблюдающиеся иногда согласованные периодические колебания численности хищников и жертв.
Для подобных моделей обычно характерен высокий уровень неустойчивости. Иными словами, при широком спектре входных параметров (таких как смертность хищников, эффективность превращения биомассы жертв в биомассу хищников и т. п.) в этих моделях рано или поздно все хищники либо вымирают, либо сначала съедают всех жертв, а потом всё равно погибают от голода.
В природных экосистемах, конечно, всё сложнее, чем в математической модели. По-видимому, существует множество факторов, способных повысить устойчивость системы «хищник–жертва», и в реальности дело редко доходит до таких резких скачков численности, как у канадских рысей и зайцев.
Экологи из Канады и США опубликовали в последнем номере журнала «Nature» статью, в которой обратили внимание на один простой и очевидный фактор, который может резко изменить поведение системы «хищник–жертва». Речь идет о групповом образе жизни.
Большинство имеющихся моделей исходят из предположения о равномерном распределении хищников и их жертв в пределах данной территории. На этом основаны расчеты частоты их встреч. Ясно, что чем выше плотность жертв, тем чаще натыкаются на них хищники. От этого зависит число нападений, в том числе успешных, и в конечном счете - интенсивность выедания жертв хищниками. Например, при избытке жертв (если не надо тратить время на поиски), скорость выедания будет ограничиваться только временем, необходимым хищнику для того, чтобы поймать, убить, съесть и переварить очередную жертву. Если добыча попадается редко, главным фактором, определяющим скорость выедания, становится время, необходимое для поисков жертвы.
В экологических моделях, используемых для описания систем «хищник–жертва», ключевую роль играет именно характер зависимости интенсивности выедания (число жертв, съедаемых одним хищником в единицу времени) от плотности популяции жертв. Последняя оценивается как число животных на единицу площади.
Следует обратить внимание, что при групповом образе жизни как жертв, так и хищников исходное допущение о равномерном пространственном распределении животных не выполняется, и поэтому все дальнейшие расчеты становятся неверными. Например, при стадном образе жизни жертв вероятность встречи с хищником фактически будет зависеть не от количества отдельных животных на квадратный километр, а от количества стад на ту же единицу площади. Если бы жертвы были распределены равномерно, хищники натыкались бы на них гораздо чаще, чем при стадном образе жизни, поскольку между стадами образуются обширные пространства, где нет никакой добычи. Сходный результат получается и при групповом образе жизни хищников. Прайд львов, бредущий по саванне, заметит ненамного больше потенциальных жертв, чем заметил бы одинокий лев, идущий тем же путем.
В течение трех лет (с 2003-го по 2007 год) ученые вели тщательные наблюдения за львами и их жертвами (прежде всего антилопами гну) на обширной территории парка Серенгети (Танзания). Плотность популяций фиксировалась ежемесячно; регулярно оценивалась также и интенсивность поедания львами различных видов копытных. И сами львы, и семь основных видов их добычи ведут групповой образ жизни. Авторы ввели в стандартные экологические формулы необходимые поправки, учитывающие это обстоятельство. Параметризация моделей проводилась на основе реальных количественных данных, полученных в ходе наблюдений. Рассматривалось 4 варианта модели: в первом групповой образ жизни хищников и жертв игнорировался, во втором учитывался только для хищников, в третьем - только для жертв, и в четвертом - для тех и других.
|
|
Как и следовало ожидать, лучше всего соответствовал реальности четвертый вариант. Он оказался к тому же и самым устойчивым. Это значит, что при широком спектре входных параметров в этой модели оказывается возможным длительное устойчивое сосуществование хищников и жертв. Данные многолетних наблюдений показывают, что в этом отношении модель тоже адекватно отражает реальность. Численности львов и их жертв в парке Серенгети довольно устойчивы, ничего похожего на периодические согласованные колебания (как в случае с рысями и зайцами) не наблюдается.
Полученные результаты показывают, что, если бы львы и антилопы гну жили поодиночке, рост численности жертв приводил бы к стремительному ускорению их выедания хищниками. Благодаря групповому образу жизни этого не происходит, активность хищников возрастает сравнительно медленно, и общий уровень выедания остается низким. По мнению авторов, подкрепленному рядом косвенных свидетельств, численность жертв в парке Серенгети лимитируется вовсе не львами, а кормовыми ресурсами.
Если выгоды коллективизма для жертв вполне очевидны, то в отношении львов вопрос остается открытым. Данное исследование наглядно показало, что групповой образ жизни для хищника имеет серьезный недостаток - по сути дела, из-за него каждому отдельному льву достается меньше добычи. Очевидно, что этот недостаток должен компенсироваться какими-то очень весомыми преимуществами. Традиционно считалось, что общественный образ жизни львов связан с охотой на крупных животных, с которыми трудно справиться в одиночку даже льву. Однако в последнее время многие специалисты (и в том числе авторы обсуждаемой статьи) стали сомневаться в правильности этого объяснения. По их мнению, коллективные действия необходимы львам только при охоте на буйволов, а с другими видами добычи львы предпочитают расправляться в одиночку.
Более правдоподобным выглядит предположение, что прайды нужны для регулирования чисто внутренних проблем, которых немало в львиной жизни. Например, у них распространен инфантицид - убийство самцами чужих детенышей. Самкам, держащимся группой, легче защищать своих детей от агрессоров. Кроме того, прайду гораздо легче, чем льву-одиночке, оборонять свой охотничий участок от соседних прайдов.
Источник : John M. Fryxell, Anna Mosser, Anthony R. E. Sinclair, Craig Packer. Group formation stabilizes predator–prey dynamics // Nature . 2007. V. 449. P. 1041–1043.
Имитационное моделирование системы "Хищник -Жертва"
Реферат >> Экономико-математическое моделирование... системы «Хищник -Жертва» Выполнил Гизятуллин Р.Р гр.МП-30 Проверил Лисовец Ю.П МОСКВА 2007г. Введение Взаимодействие ... модель взаимодействия хищников и жертв на плоскости. Упрощающие предположения. Попробуем сопоставить жертве и хищнику некоторый...
Хищник -Жертва
Реферат >> ЭкологияПриложения математической экологии является система хищник -жертва . Цикличность поведения этой системы в стационарной среде была... с помощью введения дополнительного нелинейного взаимодействия между хищником и жертвой . Полученая модель имеет на своей...
Конспект экология
Реферат >> ЭкологияФактором для жертвы . Поэтому взаимодействие «хищник –жертва» носит периодический характер и описывается системой уравнений Лотки... сдвиг значительно меньше, чем в системе «хищник –жертва» . Подобные взаимодействия наблюдаются и при бэтсовской мимикрии. ...
Модель «хищник - жертва» и макроэкономическая модель Гудвина
Рассмотрим биологическую модель «хищник - жертва», в которой один вид является кормом для другого. Эта модель, давно ставшая классической, была построена в первой половине XX в. итальянским математиком В. Вольтерра для объяснения колебаний уловов рыбы в Адриатическом море . В модели предполагается, что число хищников растет до тех пор, пока у них будет достаточно пищи, а увеличение поголовья хищников приводит к уменьшению популяции рыб-жертв. Когда последних становится мало, численность хищников уменьшается. Вследствие этого с некоторого момента начинается рост численности рыб-жертв, который через некоторое время вызывает рост популяции хищников. Цикл замыкается.
Пусть N x (t) и N 2 (t) - численности рыб-жертв и рыб-хищников в момент времени t соответственно. Предположим, что темп прироста численности жертв в условиях отсутствия хищников постоянен, т. е.
где а - положительная постоянная.
Появление хищника должно снизить темп прироста жертв. Будем считать, что это снижение линейно зависит от численности хищника: чем больше хищников, тем меньше темп прироста жертв. Тогда

где т > 0.
Поэтому для динамики численности рыб-жертв получаем:
Составим теперь уравнение, определяющее динамику популяции хищников. Предположим, что их численность в случае отсутствия жертв сокращается (из-за отсутствия корма) с постоянным темпом Ь, т. е.
![]()
Наличие жертв вызывает увеличение темпа прироста хищников. Предположим, что этот прирост линеен, т. е.
![]()
где п> 0.
Тогда для скорости роста рыб-хищников получаем уравнение:
В системе «хищник - жертва» (6.17)-(6.18) снижение скорости роста численности рыб-жертв, вызванное поеданием их хищниками, равно mN x N 2 , т. е. пропорционально числу их встреч с хищником. Увеличение скорости роста численности рыб-хищников, вызванное наличием жертв, равно nN x N 2 , т. е. тоже пропорционально числу встреч жертв и хищников.
Введем безразмерные переменные U = mN 2 /a и V = nN x /b. Динамика переменной U соответствует динамике хищников, а динамика переменной V - динамике жертв. В силу уравнений (6.17) и (6.18) изменение новых переменных определяется системой уравнений:
Предположим, что при t = 0 число особей обоих видов известно, следовательно, известны начальные значения новых переменных?/(0) = U 0 , К(0) = К 0 . Из системы уравнений (6.19) можно найти дифференциальное уравнение для ее фазовых траекторий:

Разделяя переменные этого уравнения, получим:


Рис. 6.10. Построение фазовой траектории ADCBA системы дифференциальных уравнений (6.19)
Отсюда с учетом начальных данных следует:
где константа интегрирования С = b(V Q - InV 0)/a - lnU 0 + U 0 .
На рис. 6.10 показано, как строится линия (6.20) при заданном значении С. Для этого в первой, во второй и в третьей четвертях соответственно строим графики функций х = V - InV, у = (Ь/а)х , у = = InU- U+C.
В силу равенства dx/dV = (V- 1)/У функция х = V- In К, определенная при V > 0, возрастает, если V> 1, и убывает, если V 1. В силу того, что cPx/dV 1 = 1/F 2 > 0, график функции л: = x(V) направлен выпуклостью вниз. Уравнение V= 0 задает вертикальную асимптоту. Наклонных асимптот эта функция не имеет. Поэтому график функции х = х(У) имеет вид кривой, изображенной в первой четверти рис. 6.10.
Аналогичным образом исследуется функция у = InU - U+ С, график которой на рис. 6.10 изображен в третьей четверти.
Если теперь мы разместим на рис. 6.10 во второй четверти график функции у = (Ь/а)х , то в четвертой четверти получим линию, которая связывает переменные U и V. Действительно, взяв точку V t на оси OV, вычисляем с помощью функции х = V - V соответствующее знание х х. После этого, используя функцию у = (Ь/а)х , по полученному значению х { находим у х (вторая четверть на рис. 6.10). Далее с помощью графика функции у = InU - U + С определяем соответствующие значения переменной U (на рис. 6.10 таких значений два - координаты точек М и N). Совокупность всех таких точек (V; U) образует искомую кривую. Из построения следует, что графиком зависимости (6.19) является замкнутая линия, содержащая внутри себя точку Е( 1, 1).
Напомним, что эту кривую мы получили, задав некоторые начальные значения U 0 и V 0 и вычислив по ним константу С. Взяв другие начальные значения, мы получим другую замкнутую линию, не пересекающую первую и также содержащую внутри себя точку Е( 1, 1). Это означает, что семейством траекторий системы (6.19) на фазовой плоскости (V, U) является множество замкнутых непересекающихся линий, концентрирующихся вокруг точки Е( 1, 1), а решения исходной модели U = Щ) и V = V{t) являются функциями, периодическими во времени. При этом максимум функции U = U(t) не попадает на максимум функции V = V(t ) и наоборот, т. е. колебания численности популяций около своих равновесных решений происходят в разных фазах.
На рис. 6.11 приведены четыре траектории системы дифференциальных уравнений (6.19) на фазовой плоскости OUV, отличающиеся начальными условиями. Одна из траекторий равновесная - это точка Е( 1, 1), которой соответствует решение U(t) = 1, V{t) = 1. Точки (U(t), V(t)) на других трех фазовых траекториях по мере увеличения времени смещаются по часовой стрелке.
Чтобы пояснить действие механизма изменения численности двух популяций, рассмотрим траекторию ABCDA на рис. 6.11. Как видим, на участке АВ и хищников, и жертв мало. Поэтому здесь популяция хищников сокращается из-за недостатка корма, а популяция жертв растет. На участке ВС численность жертв достигает высоких значений, что приводит к росту численности хищников. На участке СА хищников много, и это влечет сокращение численности жертв. При этом после прохождения точки D численность жертв уменьшается на столько, что численность популяции начинает уменьшаться. Цикл замыкается.
Модель «хищник - жертва» - пример структурно неустойчивой модели. Здесь малое изменение правой части одного из уравнений может привести к принципиальному изменению ее фазового портрета.

Рис. 6.11.

Рис. 6.12.
Действительно, если в уравнении динамики жертв учесть внутривидовую конкуренцию, то мы получим систему дифференциальных уравнений:
Здесь при т = 0 популяция жертв развивается согласно логическому закону.
При т Ф 0 ненулевое равновесное решение системы (6.21) при некоторых положительных значениях параметра внутривидовой конкуренции И является устойчивым фокусом, и соответствующие траектории «наматываются» на точку равновесия (рис. 6.12). Если же h = 0, то в этом случае особая точка Е( 1, 1) системы (6.19) является центром, и траектории представляют собой замкнутые линии (см. рис. 6.11).
Замечание. Обычно под моделью «хищник - жертва» понимают модель (6.19), фазовые траектории которой замкнуты. Однако модель (6.21) также является моделью «хищник - жертва», поскольку описывает взаимовлияние хищников и жертв.
Одним из первых приложений модели типа «хищник - жертва» в экономике для исследования циклично изменяющихся процессов является макроэкономическая модель Гудвина, в которой используется непрерывный подход к анализу взаимовлияния уровня занятости и ставки заработной платы.
В работе В.-Б. Занга изложен вариант модели Гудвина, в котором производительность труда и предложение труда растут с постоянными темпами прироста, а коэффициент выбытия фондов равен нулю. Эта модель формально приводит к уравнениям модели «хищник - жертва».
Ниже рассматривается модификация этой модели для случая отличного от нуля коэффициента выбытия фондов.
В модели используются следующие обозначения: L - численность трудящихся; w - средняя ставка заработной платы трудящихся; К - основные производственные фонды (капитал); Y - национальный доход; / - инвестиции; С - потребление; р - коэффициент выбытия фондов; N - предложение труда на рынке рабочей силы; т = Y/K - фондоотдача; а = Y/L - производительность труда; у = L/N - коэффициент занятости; х = C/Y - норма потребления в национальном доходе; К - приращение капитала в зависимости от инвестиций.
Выпишем уравнения модели Гудвина:

где а 0 , b, g, п, N 0 , г - положительные числа (параметры).
Уравнения (6.22) - (6.24) выражают следующее. Уравнение (6.22) - обычное уравнение динамики фондов. Уравнение (6.23) отражает рост ставки заработной платы при высоком уровне занятости (ставка заработной платы растет, если предложение труда невелико) и уменьшение ставки заработной платы при высоком уровне безработицы.
Тем самым уравнение (6.23) выражает закон Филлипса в линейной форме . Уравнения (6.24) означают экспоненциальный рост производительности труда и предложения труда. Предположим также, что С = wL, т. е. вся заработная плата тратится на потребление. Теперь можно преобразовать уравнения модели с учетом равенств:

Преобразуем уравнения (6.22)-(6.27). Имеем:
 где
где
где
![]()
Поэтому динамика переменных в модели Гудвина описывается системой дифференциальных уравнений:

которая формально совпадает с уравнениями классической модели «хищник - жертва». Это значит, что в модели Гудвина тоже возникают колебания фазовых переменных. Механизм колебательной динамики заключается здесь в следующем: при малой заработной плате w потребление низкое, инвестиции большие, и это приводит к росту производства и занятости у. Большая занятость у вызывает рост средней заработной платы w, что приводит к росту потребления и снижению инвестиций, падению производства и снижению занятости у.
Ниже гипотеза о зависимости ставки процента от уровня занятости рассмотренной модели используется при моделировании динамики однопродуктовой фирмы. Оказывается, что в этом случае при некоторых дополнительных допущениях модель фирмы обладает свойством цикличности рассмотренной выше модели «хищник - жертва».
- См.: Вольтерра В. Указ, соч.; Ризнииенко Г. Ю., Рубин А. Б. Указ. соч.
- См.: Занг В.-Б. Синергетическая экономика. М., 2000.
- См.: Пу Т. Нелинейная экономическая динамика. Ижевск, 2000; Тихонов А. Н.Математическая модель // Математическая энциклопедия. Т. 3. М., 1982. С. 574, 575.
Еще в 20-х гг. А. Лотка (Lotka), а несколько позднее независимо от него В. Вольтерра предложили математические модели, описывающие сопряженные колебания численности популяций хищника и жертвы. Рассмотрим самый простой вариант модели Лотки-Вольтерра. В основе модели лежит ряд допущений:
1) популяция жертв в отсутствие хищника растет экспоненциально,
2) пресс хищников тормозит этот рост,
3) смертность жертв пропорциональна частоте встреч хищника и жертвы (или иначе, пропорциональна произведению плотностей их популяций);
4) рождаемость хищника зависит от интенсивности потребления жертв.
Мгновенная скорость изменения численности популяции жертв может быть выражена уравнением
dN ж /dt = r 1 N ж - p 1 N ж N х,
где r 1 - удельная мгновенная скорость популяционного роста жертвы, p 1 - константа, связывающая смертность жертв с плотностью хищника, a N ж и N х - плотности соответственно жертвы и хищника.
Мгновенная скорость роста популяции хищника в этой модели принимается равной разности рождаемости и постоянной смертности:
dN х /dt = p 2 N ж N х – d 2 N х,
где p 2 - константа, связывающая рождаемость в популяции хищника с плотностью жертв, a d 2 - удельная смертность хищника.
Согласно приведенным уравнениям каждая из взаимодействующих популяций в своем увеличении ограничена только другой популяцией, т.е. рост числа жертв лимитируется прессом хищников, а рост числа хищников - недостаточным количеством жертв. Никакого самоограничения популяций не предполагается. Считается, например, что пищи для жертвы всегда достаточно. Также не предполагается и выхода популяции жертв из-под контроля хищника, хотя на самом деле такое бывает достаточно часто.
Несмотря на всю условность модели Лотки-Вольтерра, она заслуживает внимания уже хотя бы потому, что показывает, как даже такая идеализированная система взаимодействия двух популяций может порождать достаточно сложную динамику их численности. Решение системы этих уравнений позволяет сформулировать условия поддержания постоянной (равновесной) численности каждого из видoв. Популяция жертв сохраняет постоянную численность, если плотность хищника равна r 1 /p 1, а для того чтобы постоянство сохраняла популяция хищника, плотность жертв должна быть равна d 2 /p 2 . Если на графике отложить по оси абсцисс плотность жертв N ж , а по оси ординат - плотность хищника N х , то изоклины, показывающие условие постоянства хищника и жертвы, будут представлять собой две прямые, перпендикулярные друг другу и координатным осям (рис. 6,а). При этом предполагается, что ниже определенной (равной d 2 /p 2) плотности жертв плотность хищника всегда будет уменьшаться, а выше - всегда увеличиваться. Соответственно и плотность жертвы возрастает, если плотность хищника ниже значения, равного r 1 /p 1 , и уменьшается, если, она выше этого значения. Точка пересечения изоклин соответствует условию постоянства численности хищника и жертвы, а другие точки на плоскости этого графика совершают движение по замкнутым траекториям, отражая, таким образом, регулярные колебания численности хищника и жертвы (рис. 6,б). Размах колебаний определяется начальным соотношением плотностей хищника и жертвы. Чем ближе оно к точке пересечения изоклин, тем меньше окружность, описываемая векторами, и соответственно меньше амплитуда колебаний.
Рис. 6. Графическое выражение модели Лотки-Вольтера для системы хищник-жертва.
Одна из первых попыток получения колебаний численности хищника и жертвы в лабораторных экспериментах принадлежала Г.Ф. Гаузе (Gause). Объектами этих экспериментов были инфузория парамеция (Paramecium caudatum ) и хищная инфузория дидиниум (Didinium nasutum ). Пищей для парамеции служила регулярно вносимая в среду взвесь бактерий, а дидиниум питался только парамециями. Данная система оказалась крайне неустойчивой: пресс хищника по мере увеличения его численности приводил к полному истреблению жертв, после чего вымирала и популяция самого хищника. Усложняя опыты, Гаузе устраивал убежище для жертвы, внося в пробирки с инфузориями немного стеклянной ваты. Среди нитей ваты могли свободно перемещаться парамеции, но не могли дидиниумы. В таком варианте опыта дидиниум съедал всех парамеций, плавающих в свободной от ваты части пробирки, и вымирал, а популяция парамеции затем восстанавливалась за счет размножения особей, уцелевших в убежище. Некоторого подобия колебаний численности хищника и жертвы Гаузе удалось добиться только в том случае, когда он время от времени вносил в культуру и жертву и хищника, имитируя таким образом иммиграцию.
Через 40 лет после работы Гаузе его опыты были повторены Л. Лакинбиялом (Luckinbill), использовавшим в качестве жертвы инфузорию Paramecium aurelia , а в качестве хищника того же Didinium nasutum . Лакинбиллу удалось получить несколько циклов колебаний численности этих популяций, но только в том случае, когда плотность парамеций была лимитирована нехваткой пищи (бактерий), а в культуральную жидкость добавляли метилцеллюлозу - вещество, снижающее скорость движения как хищника, так и жертвы и потому уменьшающее частоту их возможных встреч. Оказалось также, что добиться колебаний хищника и жертвы легче, если увеличить объем экспериментального сосуда, хотя условие пищевого лимитирования жертвы и в этом случае обязательно. Если же к системе сосуществующих в колебательном режиме хищника и жертвы добавляли избыточную пищу, то ответом был, быстрый рост численности жертвы, за которым следовало возрастание численности хищника, приводящее в свою очередь к полному истреблению популяции жертвы.
Модели Лотки и Вольтерры послужили толчком для разработки ряда других более реалистичных моделей системы хищник-жертва. В частности, довольно простая графическая модель, анализирующая соотношение разных изоклин жертвы хищника, была предложена М. Розенцвейгом и Р. Мак-Артуров (Rosenzweig, MacArthur). Согласно этим авторам, стационарная (= постоянная) численность жертвы в координатных осях плотности хищника и жертвы может быть представлена в виде выпуклой изоклины (рис. 7,а). Одна точка пересечения изоклины сосью плотности жертвы соответствует минимальной допустимой плотности жертвы (нижеее популяция подвержена очень большому риску вымирания хотя бы из-за малой частоты встреч самцов и самок), а другая - максимальной, определяемой количеством имеющейся пищи или поведенческими особенностями самой жертвы. Подчеркнем, что речь идет пока о минимальной и максимальной плотностях в отсутствие хищника. При появлении хищника и увеличении его численности минимальная допустимая плотность жертвы, очевидно, должна быть выше, а максимальная - ниже. Каждому значению плотности жертвы должна соответствовать некоторая плотность хищника, при которой достигается постоянство популяции жертвы. Геометрическое место таких точек и есть изоклина жертвы в координатах плотности хищника и жертвы. Векторы, показывающие направление изменения плотности жертвы (ориентированные горизонтально), имеют разную направленность по разные стороны от изоклины (рис. 7,а).

Рис. 7. Изоклины стационарных популяций жертвы (а) и хищника (б).
Для хищника в тех же координатах также построена изоклина, отвечающая стационарному состоянию его популяции. Векторы, показывающие направление изменения численности хищника, ориентированы вверх или вниз в зависимости от того, по какую сторону от изоклины они находятся. Форма изоклины хищника, показанная на рис. 7,б. определяется, во-первых, наличием некоторой минимальной плотности жертвы, достаточной для поддержания популяции хищника (при более низкой плотности жертвы хищник не может увеличивать свою численность), а во-вторых, наличием некоторой максимальной плотности самого хищника, при превышении которой численность будет снижаться независимо от обилия жертв.

Рис. 8. Возникновение колебательных режимов в системе хищник-жертва в зависимости от расположения изоклин хищника и жертвы.
При совмещении изоклин жертвы и хищника на одном, графике возможны три различных варианта (рис. 8). Если изоклина хищника пересекает изоклину жертвы в том месте, где она уже снижается (при высокой плотности жертв), векторы, показывающие изменение численности хищника и жертвы, образуют траекторию, закручивающуюся во внутрь, что соответствует затухающим колебаниям численности жертвы и хищника (рис. 8,а). В том случае, когда изоклина хищника пересекает изоклину жертвы в ее восходящей части (т.е. в области низких значений плотности жертв), векторы образуют раскручивающуюся траекторию, а колебания численности хищника и жертвы происходят соответственно с возрастающей амплитудой (рис. 8,б). Если же изоклина хищника пересекает изоклину жертвы в области ее вершины, то векторы образуют замкнутый круг, а колебания численности жертвы и хищника характеризуются стабильной амплитудой и периодом (рис. 8,в).
Иными словами, затухающие колебания соответствуют ситуации, при которой хищник ощутимо воздействует на популяцию жертв, достигнувшую только очень высокой плотности (близкой к предельной), а колебания возрастающей амплитуды возникают, тогда, когда хищник способен быстро увеличивать свою численность даже при невысокой плотности жертв и таким образом быстро ее уничтожить. В других вариантах своей модели Poзенцвейг и Мак-Артур показали, что стабилизировать колебания хищник-жертва можно, введя «убежище», т.е. предположив, что в области низкой плотности жертв существует область, где численность жертвы растет независимо от количества имеющихся хищников.
Стремление сделать модели более реалистичными путем их усложнения проявилось в работах не только теоретиков, но и экспериментаторов. В частности, интересные результаты были получены Хаффейкером (Huffaker), показавшим возможность сосуществования хищника и жертвы в колебательном режиме на примере мелкого растительноядного клеща Eotetranychus sexmaculatus и нападающего на него хищного клеща Typhlodromus occidentalis . В качестве пищи для растительноядного клеща использовали апельсины, помещенные на подносы с лунками (вроде тех, что используются для хранения и перевозки яиц). В первоначальном варианте на одном подносе было 40 лунок, причем в некоторых из них находились апельсины (частично очищенные от кожуры), а в других - резиновые мячики. Оба вида клещей размножаются партеногенетически очень быстро, и поэтому характер их популяционной динамики можно выявить за сравнительно короткий срок. Поместив на поднос 20 самок растительноядного клеща Хаффейкер наблюдал быстрый рост его популяции, которая стабилизировалась на уровне 5-8 тыс. особей (в расчете на один апельсин). Если к растущей популяции жертвы добавляли несколько особей хищника, то популяция последнего быстро увеличивала свою численность и вымирала, когда все жертвы оказывались съеденными.
Увеличив размер подноса до 120 лунок, в которых отдельные апельсины были случайно разбросаны среди множества резиновых мячиков, Хаффейкеру удалось продлить сосуществование хищника и жертвы. Важную роль во взаимодействии хищника и жертвы, как выяснилось, играет соотношение скоростей их расселения. Хаффейкер предположил, что, облегчив передвижение жертвы и затруднив передвижение хищника, можно увеличить время их сосуществования. Для этого на подносе из 120 лунок среди резиновых мячиков располагали случайным образом 6 апельсинов, причем вокруг лунок с апельсинами были устроены преграды из вазелина, препятствовавшие расселению хищника, а для облегчения расселения жертвы на подносе были укреплены деревянные колышки, служившие своего рода «взлетными площадками» для растительноядных клещей (дело в том, что этот вид выпускает тонкие нити и с помощью их может парить в воздухе, распространяясь по ветру). В таком усложненном местообитании хищник и жертва сосуществовали в течение 8 месяцев, продемонстрировав три полных цикла колебаний численности. Наиболее важные условия этого сосуществования следующие: гетерогенность местообитания (в смысле наличия в ней пригодных и непригодных для обитания жертвы участков), а также возможность миграции жертвы и хищника (с сохранением некоторого преимущества жертвы в скорости этого процесса). Иными словами, хищник может полностью истребить то или иное локальное скопление жертв, но часть особей жертвы успеет мигрировать и дать начало другим локальным скоплениям. До новых локальных скоплений хищник рано или поздно тоже доберется, но тем временем жертва успеет расселиться в другие места (в том числе и в те, где она обитала раньше, но потом была истреблена).
Нечто подобное тому, что наблюдал Хаффейкер в эксперименте, встречается и в природных условиях. Так, например, бабочка кактусовая огневка (Cactoblastis cactorum ), завезенная в Австралию, значительно снизила численность кактуса опунции, но не уничтожила его полностью именно потому, что кактус успевает расселиться немного быстрее. В тех местах, где опунция истребляется полностью, перестает встречаться и огневка. Поэтому, когда через некоторое время сюда вновь проникает опунция, то в течение определенного периода она может произрастать без риска быть уничтоженной огневкой. Со временем, однако, огневка снова здесь появляется и, быстро размножаясь, уничтожает опунцию.
Говоря о колебаниях хищник-жертва, нельзя не упомянуть и о циклических изменениях численности зайца и рыси в Канаде, прослеженных по материалам статистики заготовок пушнины компанией Гудзон-Бэй с конца XVIII вплоть до начала XX в. Этот пример нередко рассматривался как классическая иллюстрация колебаний хищник-жертва, хотя на самом деле мы видим только следование роста численности популяции хищника (рыси) за ростом численности жертвы (зайца). Что же касается снижения численности зайцев после каждого подъема, то оно не могло объясняться только возросшим прессом хищников, а было связано с другими факторами, по-видимому, прежде всего нехваткой корма в зимний период. К такому выводу пришел, в частности, М. Джилпин (Gilpin), пытавшийся проверить, могут ли быть описаны эти данные классической моделью Лотки-Вольтерры. Результаты проверки показали, что удовлетворительного соответствия модели нет, но как ни странно, оно становилось лучше, если хищника и жертву меняли местами, т.е. трактовали рысь как «жертву», а зайца - как «хищника». Подобная ситуация нашла свое отражение и в шутливом названии статьи («Едят ли зайцы рысей?»), по сути своей очень серьезной и опубликованной в серьезном научном журнале.
Федеральное агентство по образованию
Государственное образовательное учреждение
высшего профессионального образования
«Ижевский государственный технический университет»
Факультет «Прикладная математика»
Кафедра «Математическое моделирование процессов и технологий»
Курсовая работа
по дисциплине «Дифференциальные уравнения»
Тема: «Качественное исследование модели хищник-жертва»
Ижевск 2010
ВВЕДЕНИЕ
1. ПАРАМЕТРЫ И ОСНОВНОЕ УРАВНЕНИЕ МОДЕЛИ «ХИЩНИК-ЖЕРТВА»
2.2 Обобщенные модели Вольтера типа «хищник-жертва».
3. ПРАКТИЧЕСКОЕ ПРИМЕНЕНИЕ МОДЕЛИ «ХИЩНИК-ЖЕРТВА»
ЗАКЛЮЧЕНИЕ
СПИСОК ЛИТЕРАТУРЫ
ВВЕДЕНИЕ
В настоящее время задачи экологии имеют первостепенное значение. Важным этапом решения этих задач является разработка математических моделей экологических систем.
Одной из основных задач экологии па современном этапе является изучение структуры и функционирования природных систем, поиск общих закономерностей. Большое влияние на экологию оказала математика, способствующая становлению математической экологии, особенно такие её разделы, как теория дифференциальных уравнений, теория устойчивости и теория оптимального управления.
Одной из первых работ в области математической экологии была работа А.Д. Лотки (1880 - 1949), который первый описал взаимодействие различных популяций, связанных отношениями хищник - жертва. Большой вклад в исследование модели хищник -жертва внесли В. Вольтерра (1860 - 1940), В.А. Костицин (1883-1963) В настоящее время уравнения описывающие взаимодействие популяций, называются уравнениями Лотки - Вольтерра.
Уравнения Лотки - Вольтерра описывают динамику средних величин - численности популяции. В настоящее время на их основе построены более общие модели взаимодействия популяций, описываемые интегро-дифференциальными уравнениями, исследуются управляемые модели хищник - жертва.
Одной из важных проблем математической экологии является проблема устойчивости экосистем, управления этими системами. Управление может осуществляться с целью перевода системы из одного устойчивого состояния в другое, с целью её использования или восстановления.
1. ПАРАМЕТРЫ И ОСНОВНОЕ УРАВНЕНИЕ МОДЕЛИ ХИЩНИК-ЖЕРТВА
Попытки математического моделирования динамики как отдельных биологических популяций, так и сообществ, включающих взаимодействующие популяции различных видов, предпринимались давно. Одна из первых моделей роста изолированной популяции (2.1) была предложена еще в 1798 г. Томасом Мальтусом:
Данная модель задается следующими параметрами:
N - численность популяции;
Разность между коэффициентами рождаемости и смертности.
Интегрируя это уравнение получаем:
![]() , (1.2)
, (1.2)
где N(0) – численность популяции в момент t = 0. Очевидно, что модель Мальтуса при > 0 дает бесконечный рост численности, что никогда не наблюдается в природных популяциях, где ресурсы, обеспечивающие этот рост, всегда ограничены. Изменения численности популяций растительного и животного мира нельзя описывать простым законом Мальтуса, на динамику роста влияют многие взаимосвязанные причины – в частности, размножение каждого вида саморегулируется и видоизменяется так, чтобы этот вид сохранялся в процессе эволюции.
Математическим описанием этих закономерностей занимается математическая экология – наука об отношениях растительных и животных организмов и образуемых ими сообществ между собой и с окружающей средой.
Наиболее серьезное исследование моделей биологических сообществ, включающих в себя несколько популяций различных видов, было проведено итальянским математиком Вито Вольтерра:
 ,
,
где - численность популяции;
Коэффициенты естественного прироста (или смертности) популяции; - коэффициенты межвидового взаимодействия. В зависимости от выбора коэффициентов модель описывает либо борьбу видов за общий ресурс, либо взаимодействие типа хищник - жертва, когда один вид является пищей для другого. Если в работах других авторов основное внимание уделялось построению различных моделей, то В. Вольтерра провел глубокое исследование построенных моделей биологических сообществ. Именно с книги В. Вольтерра, по мнению многих ученых, началась современная математическая экология.
2. КАЧЕСТВЕННОЕ ИССЛЕДОВАНИЕ ЭЛЕМЕНТАРНОЙ МОДЕЛИ «ХИЩНИК-ЖЕРТВА»
2.1 Модель трофического взаимодействия по типу «хищник-жертва»
Рассмотрим модель трофического взаимодействия по типу «хищник-жертва», построенную В. Вольтерром. Пусть имеется система, состоящая из двух видов, из которых один поедает другой.
Рассмотрим случай, когда один из видов является хищником, а другой - жертвой, и будем считать, что хищник питается только жертвой. Примем следующую простую гипотезу:
Коэффициент прироста жертвы;
Коэффициент прироста хищника;
Численность популяции жертвы;
Численность популяции хищника;
Коэффициент естественного прироста жертвы;
Скорость потребления жертвы хищником;
Коэффициент смертности хищника в отсутствие жертвы;
Коэффициент «переработки» хищником биомассы жертвы в собственную биомассу.
Тогда динамика численности популяций в системе хищник - жертва будет описываться системой дифференциальных уравнений (2.1):
 (2.1)
(2.1)
где все коэффициенты положительные и постоянные.
Модель имеет равновесное решение (2.2):
По модели (2.1) доля хищников в общей массе животных выражается формулой (2.3):
 (2.3)
(2.3)
Анализ устойчивости состояния равновесия по отношению к малым возмущениям показал, что особая точка (2.2) является «нейтрально» устойчивой (типа «центр»), т. е. любые отклонения от равновесия не затухают, но переводят систему в колебательный режим с амплитудой, зависящей от величины возмущения. Траектории системы на фазовой плоскости имеют вид замкнутых кривых, расположенных на различных расстояниях от точки равновесия (рис. 1).

Рис. 1 – Фазовый «портрет» классической вольтерровой системы «хищник-жертва»
Разделив первое уравнение системы (2.1) на второе, получим дифференциальное уравнение (2.4) для кривой на фазовой плоскости .
 (2.4)
(2.4)
Интегрируя данное уравнение получим:
![]() (2.5)
(2.5)
где - постоянная интегрирования, где 
Несложно показать, что движение точки по фазовой плоскости будет происходить только в одну сторону. Для этого удобно сделать замену функций и , перенеся начало координат на плоскости в стационарную точку (2.2) и введя затем полярные координаты:
 (2.6)
(2.6)
В таком случае, подставив значения системы (2.6) в систему (2.1), будем иметь:
 (2.7)
(2.7)
Умножив первое уравнение на , а второе - на и сложив их, получим:
После аналогичных алгебраических преобразований получим уравнение для :
Величина , как видно из (4.9), всегда больше нуля. Таким образом, не меняет знака, и вращение все время идет в одну сторону.
Интегрируя (2.9) найдем период:
Когда мало, то уравнения (2.8) и (2.9) переходят в уравнения эллипса. Период обращения в этом случае равен:
 (2.11)
(2.11)
Исходя из периодичности решений уравнений (2.1), можно получить некоторые следствия. Представим для этого (2.1) в виде:
 (2.12)
(2.12)
и проинтегрируем по периоду:
 (2.13)
(2.13)
Так как подстановки от и в силу периодичности обращаются в нуль, средние по периоду оказываются равными стационарным состояниям (2.14):
 (2.14)
(2.14)
Простейшие уравнения модели «хищник-жертва» (2.1) обладают рядом существенных недостатков. Так, в них предполагается неограниченность пищевых ресурсов для жертвы и неограниченный рост хищника, что противоречит экспериментальным данным. Кроме того, как видно из рис. 1, ни одна из фазовых кривых не выделена с точки зрения устойчивости. При наличии даже небольших возмущающих воздействий траектория системы будет все дальше уходить от положения равновесия, амплитуда колебаний расти, и система достаточно быстро разрушится.
Несмотря на недостатки модели (2.1), представления о принципиально колебательном характере динамики системы «хищник- жертва» получили широкое распространение в экологии. Взаимодействиями «хищник-жертва» объясняли такие явления, как колебания численности хищных и мирных животных в промысловых зонах, колебания в популяциях рыб, насекомых и т. д. На самом деле колебания численности могут быть обусловлены и другими причинами.
Предположим, что в системе хищник - жертва происходит искусственное уничтожение особей обоих видов, и рассмотрим вопрос о том, каким образом уничтожение особей влияет на средние значения их численности, если осуществляется пропорционально этой численности с коэффициентами пропорциональности и соответственно для жертвы и хищника. С учетом сделанных предположений систему уравнений (2.1) перепишем в виде:
 (2.15)
(2.15)
Предположим, что , т. е. коэффициент истребления жертвы меньше коэффициента ее естественного прироста. В этом случае также будут наблюдаться периодические колебания численности. Вычислим средние значения численностей:
 (2.16)
(2.16)
Таким образом, если , то средняя численность популяций жертвы возрастает, а хищника - убывает.
Рассмотрим случай, когда коэффициент истребления жертвы больше коэффициента ее естественного прироста, т. Е . В этом случае ![]() при любых , и, следовательно, решение первого уравнения (2.15) ограничено сверху экспоненциально убывающей функцией
при любых , и, следовательно, решение первого уравнения (2.15) ограничено сверху экспоненциально убывающей функцией ![]() , т. е. при .
, т. е. при .
Начиная с некоторого момента времени t, при котором , решение второго уравнения (2.15) также начинает убывать и при стремится к нулю. Таким образом, в случае оба вида исчезают.
2.1 Обобщенные модели Вольтера типа «хищник-жертва»
Первые модели В. Вольтерра, естественно, не могли отражать все стороны взаимодействия в системе хищник - жертва, поскольку они были в значительной мере упрощены относительно реальных условий. Например, если численность хищника равна нулю, то из уравнений (1.4) следует, что численность жертвы неограниченно возрастает, что не соответствует действительности. Однако ценность этих моделей состоит именно в том, что они были основой, на которой быстрыми темпами начала развиваться математическая экология.
Появилось большое число исследований различных модификаций системы хищник - жертва, где были построены более общие модели, учитывающие в той или иной степени реальную ситуацию в природе.
В 1936 г. А.Н. Колмогоров предложил использовать для описания динамики системы хищник - жертва следующую систему уравнении:
 , (2.17)
, (2.17)
где убывает с возрастанием численности хищников, а возрастает с увеличением численности жертвы.
Эта система дифференциальных уравнений в силу ее достаточной общности позволяет хорошо учитывать реальное поведение популяций и вместе с тем проводить качественный анализ ее решений.
Позднее в своей работе, Колмогоров исследовал подробно менее общую модель:
 (2.18)
(2.18)
Различные частные случаи системы дифференциальных уравнений (2.18) исследовались многими авторами. В таблице приведены различные частные случаи функций , , .
Таблица 1 - Различные модели сообщества «хищник-жертва»
| Авторы | |||
| Вольтерра-Лотка | |||
| Гаузе | |||
| Пислоу | |||
| Холинг | |||
| Ивлев | |||
| Рояма | |||
| Шимазу | |||
| Мэй |
математическое моделирование хищник жертва
3. ПРАКТИЧЕСКОЕ ПРИМЕНЕНИЕ МОДЕЛИ ХИЩНИК-ЖЕРТВА
Рассмотрим математическую модель совместного существования двух биологических видов (популяций) типа "хищник - жертва", называемую моделью Вольтерра - Лотки.
Пусть два биологических вида совместно обитают в изолированной среде. Среда стационарна и обеспечивает в неограниченном количестве всем необходимым для жизни один из видов, который будем называть жертвой. Другой вид - хищник также находится в стационарных условиях, но питается лишь особями первого вида. Это могут быть караси и щуки, зайцы и волки, мыши и лисы, микробы и антитела и т. д. Будем для определенности называть их карасями и щуками.
Заданы следующие начальные показатели:
Со временем число карасей и щук меняется, но так как рыбы в пруду много, то не будем различать 1020 карасей или 1021 и поэтому будем считать и непрерывными функциями времени t. Будем называть пару чисел (,) состоянием модели.
Очевидно, что характер изменения состояния (,) определяется значениями параметров. Изменяя параметры и решая систему уравнений модели, можно исследовать закономерности изменения состояния экологической системы во времени.
В экосистеме скорость изменения численности каждого вида также будем считать пропорциональной его численности, но только с коэффициентом, который зависит от численности особей другого вида. Так, для карасей этот коэффициент уменьшается с увеличением числа щук, а для щук увеличивается с увеличением числа карасей. Будем считать эту зависимость также линейной. Тогда получим систему из двух дифференциальных уравнений:

Эта система уравнений и называется моделью Вольтерра-Лотки. Числовые коэффициенты , , - называются параметрами модели. Очевидно, что характер изменения состояния (,) определяется значениями параметров. Изменяя эти параметры и решая систему уравнений модели, можно исследовать закономерности изменения состояния экологической системы.
Проинтегрируем оба уравнения систему по t, которое будет изменяться от - начального момента времени, до , где T – период, за который происходят изменения в экосистеме. Пусть в нашем случае период равен 1 году. Тогда система принимает следующий вид:
 ;
;
 ;
;
Принимая = и = приведем подобные слагаемые, получим систему, состоящую из двух уравнений:

Подставив в полученную систему исходные данные получим популяцию щук и карасей в озере спустя год:





